
資訊中心
作者:香雪生命科學(xué)-王定國
發(fā)布時(shí)間:2023-02-23
閱讀:10662
12世紀最杰出的哲學(xué)家彼得·阿貝拉德(Peter Abélard)曾經(jīng)說(shuō)過(guò):“The beginning of wisdom is found in doubting; by doubting we come to question, and by seeking we may come upon the truth.”智慧的開(kāi)始源自懷疑;通過(guò)懷疑,我們質(zhì)疑問(wèn)題,通過(guò)尋求探索,我們可以找到真理。本篇文獻翻譯(節選[1]), 我們來(lái)深入了解一下與CD8αβ截然不同的CD8αα,正是因為對CD8αα的不斷懷疑,科學(xué)家對CD8αα有了更多的認識。
摘要:CD8是一種表達在造血細胞上的糖蛋白。CD8的兩種亞型,即CD8αβ和CD8αα,已經(jīng)被證實(shí)在表達和功能上是不同的。CD8αβ作為T(mén)CR的輔助受體,增強其功能親和力(avidity),并組成性地表達于MHC I類(lèi)限制性的T細胞上,而表達CD8αα的T細胞,則與傳統胸腺選擇的、MHC限制的CD4+或CD8αβ+ T細胞不同。與CD8αβ輔助受體功能不一致的是,CD8αα降低TCR的抗原敏感性,并且它可以瞬時(shí)或者永久地表達于T細胞上,無(wú)論該T細胞的MHC限制性或者是否存在傳統輔助受體。也就是說(shuō),CD8αα可以表達在CD4+或者CD8+ T細胞上,也可以和CD8αβ同時(shí)表達在同一T細胞上。這些發(fā)現表明,T細胞上的CD8αα標志著(zhù)一個(gè)特殊的分化階段,它很可能作為T(mén)CR的共抑制受體,負向調節T細胞的活化。
前言:CD8αα在T細胞上表達時(shí),經(jīng)常被認為是一種無(wú)效的TCR輔助受體。與傳統的TCR輔助受體CD4和CD8αβ類(lèi)似,CD8αα也結合MHC配體,并被作為譜系決定因子,以鑒定在多個(gè)方面(如:起源、個(gè)體發(fā)生、特異性、功能)偏離主流T細胞亞群的T細胞。在小鼠中,CD8αα單陽(yáng)性(SP)T細胞在小腸上皮內淋巴細胞(IEL)中占主導地位,并且通常顯示出活化的表型,同時(shí)具有先天性的特征。此外,CD8αα和CD8αβ在胸腺細胞上的調節是靈活的,取決于其發(fā)育階段。CD8αα也可以與CD4或CD8αβ一起表達在活化的成熟TCRαβ+ T細胞上,這意味著(zhù)CD8αα可能不是作為一種傳統的與MHC I類(lèi)結合的TCR輔助受體。
表達CD8αα的成熟T細胞的激活狀態(tài)表明CD8αα與TCR-CD3復合物之間存在關(guān)系。CD8αα在TCRγδ+以及TCRαβ+細胞上表達,或與CD4、CD8αβ一起在MHC II類(lèi)或MHC I類(lèi)限制的T細胞上表達,表明這種關(guān)系不受TCR或輔助受體性質(zhì)的限制,而且與MHC的限制性也無(wú)關(guān)。這些發(fā)現也暗示了,CD8αα和TCR-CD3激活復合體之間的聯(lián)系不是由TCR介導的,而是與CD3復合體信號模塊之間存在功能上的聯(lián)系。CD8α胞質(zhì)尾與早期Src激酶p56lck和T細胞激活連接體(LAT)相結合,將CD8αα連接到近端和遠端的TCR-CD3激活信號傳導級聯(lián)上,其中,p56lck的作用是對CD3上的“免疫受體酪氨酸激活基序”(ITAMs)進(jìn)行磷酸化,LAT的作用是介導進(jìn)一步的下游信號傳導。盡管有這種結合,且盡管CD8αα與MHC I類(lèi)配體有相互作用的能力,但CD8αα既不支持對MHC I類(lèi)限制性胸腺細胞的正向選擇,也不能有效地促進(jìn)CD8依賴(lài)的MHC I類(lèi)限制性的TCR的激活。進(jìn)一步地,當與CD8αβ一起表達時(shí),CD8αα可能會(huì )降低而不是增強CD8αβ-TCR:Ag-MHC激活復合物的功能親和力,這意味著(zhù)CD8αα對TCR-CD3激活復合物的作用是抑制性的。CD8αα的差異性、獨立性、高度調控的表達以及其獨特的生物學(xué)特點(diǎn)表明CD8αα不是CD8αβ的功能同源物,卻表明CD8αα可能作為一種有效的TCR共抑制受體,而不是功能性的TCR輔助受體。
CD8αα的抑制受體功能: 歷史上,CD8αα一直被用來(lái)作為研究CD8功能的方便分子。通過(guò)CD8α基因的轉導,表明CD8αα可以作為一個(gè)粘附分子發(fā)揮作用,能夠與未被TCR結合的MHC I類(lèi)配體相互作用而加強整體的親和力。無(wú)尾的CD8αα可以在同樣程度上增強反應性,這表明CD8αα的粘附作用完全取決于其胞外部分與MHC I類(lèi)分子的相互作用。然而,細胞-細胞間的結合試驗或可溶性分子的SPR分析表明,與CD8αα相比,CD8αβ并沒(méi)有增加細胞粘附性或對MHC I類(lèi)配體的親和力,這與CD8αβ TCR 輔助受體的增強功能不同。這種由CD8αα或CD8αβ介導的類(lèi)似粘附作用,與更高級的膜結合形式的CD8αβ作為T(mén)CR輔助受體的功能是截然不同的。因此,強調CD8β作為有效的CD8 TCR輔助受體標志的重要性,也相當于支持CD8αα不是CD8αβ TCR 輔助受體功能同源物的觀(guān)點(diǎn)。當TCR與弱抗原結合時(shí),需要有高效的CD8αβ輔助受體功能而不是增加粘附性,這一現象意味著(zhù)TCR輔助受體的功能是增強低親和力TCR的抗原敏感性。因此,具有低親和力TCR的T細胞在激活和初始選擇時(shí)依賴(lài)于輔助受體。考慮到這一點(diǎn),一項用逆轉錄病毒將CD8依賴(lài)性和非依賴(lài)性TCR(也即低親和力和高親和力TCR)分別轉導到從野生型或CD8β缺陷型小鼠分離的原代T細胞的研究表明,CD8αα不能支持CD8依賴(lài)性TCR(即低親和力TCR)的激活,進(jìn)而說(shuō)明CD8αα不具有TCR輔助受體功能。相反,在沒(méi)有輔助受體的情況下依然發(fā)揮作用的高親和力TCR出現在雙陰性(DN)T細胞上,這些T細胞卻經(jīng)常表達CD8αα。CD8αα與高親和力的TCR一起表達,或者是通過(guò)增加TCR信號強度對CD8αα的誘導,都進(jìn)一步使人們懷疑CD8αα作為一個(gè)輔助受體來(lái)提高TCR抗原敏感性的功能。在雙陰性(DN)胸腺細胞上強制轉基因表達CD8αα大大削弱了細胞內鈣反應,并阻斷了對TCR應答反應時(shí)信號通路組分的有效酪氨酸磷酸化,這表明CD8αα可能作為T(mén)CR的一個(gè)負調控器發(fā)揮作用。以此一致的是,用TCR的轉基因細胞系(表達相同親和力的TCR)已經(jīng)證明了,當與CD8αβ共表達時(shí),CD8αα能夠特異性地抑制由CD8αβ介導增加的抗原敏感性。
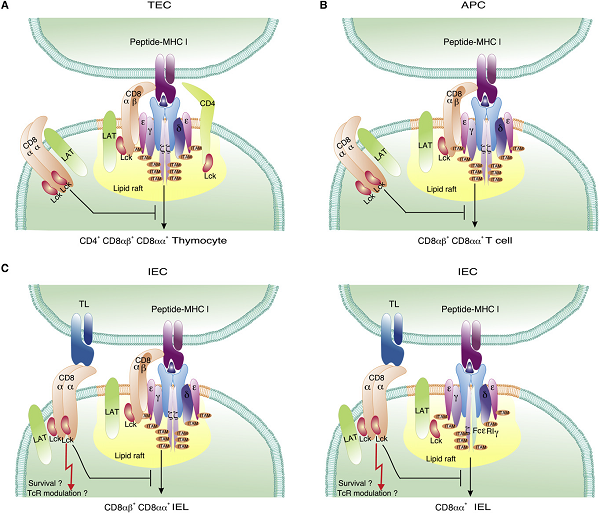
圖1.CD8αα共抑制受體在不同細胞類(lèi)型上的功能
與傳統的輔助受體CD8αβ和CD4不同,CD8αα抑制受體被排除在脂質(zhì)筏之外,脂質(zhì)筏含有與抗原呈遞MHC分子相結合的TCR-CD3(CD3ζ 和/或 FcεRγ+)激活復合物。CD8αα共抑制受體通過(guò)破壞脂質(zhì)筏和封閉TCR啟動(dòng)下游信號傳導所需的信號分子來(lái)負向調節TCR的激活。
(A)CD4+CD8αβ+CD8αα+ 三陽(yáng)的胸腺細胞在胸腺激動(dòng)劑選擇期間與MHC和自身抗原相互作用。CD8αα抑制受體可能封閉Lck和LAT,使得從激動(dòng)劑選擇TCR接收到的信號強度暫時(shí)性降低。
(B) 一些外周的CD8αβ+T細胞在被激動(dòng)劑配體初級激活后會(huì )短暫地誘導CD8αα。這些激活的T細胞上的CD8αα抑制受體可能臨時(shí)性地封閉Lck和LAT,使得通過(guò)激動(dòng)劑觸發(fā)的TCR所接收的信號強度暫時(shí)減少。
(C) 左圖:傳統的輔助受體依賴(lài)的效應T細胞在腸道微環(huán)境的條件下重新產(chǎn)生CD8αα。CD8αα抑制受體的組成性存在可能會(huì )提高這些位于抗原豐富的腸道環(huán)境中的有抗原經(jīng)歷的T細胞的激活閾值。CD8αα可能與它的配體TL相互作用,TL由腸道上皮細胞大量表達(其α3上有氨基酸突變,不能遞呈抗原肽),并促進(jìn)有抗原經(jīng)歷T細胞的長(cháng)期存活。右圖DN TCRαβ+胸腺細胞遷移到腸道后重新產(chǎn)生CD8αα。在這些自身特異性輔助受體非依賴(lài)性T細胞上CD8αα抑制受體組成性地表達可能阻止異常的自我反應。CD8αα與TL配體的相互作用可以促進(jìn)這些有自身抗原經(jīng)歷T細胞的長(cháng)期存活。
與包括CTLA-4在內的其他TCR共抑制受體類(lèi)似,CD8αα可以通過(guò)TCR-CD3復合物在激活時(shí)被誘導,誘導程度與信號強度正相關(guān)。激活誘導的CD8αα與激活復合體的功能親和力直接相關(guān),而與所參與的TCR對CD8αβ輔助受體的依賴(lài)性成負相關(guān)。因為CD8αα的共同表達有效地降低了TCR的功能親和力,并明顯減少或完全消除了激活,所以可以得出結論,CD8αα不是一個(gè)冗余的輔助受體,而是更有可能作為一個(gè)有效的TCR共抑制受體來(lái)發(fā)揮作用。由于CD8αα可以在已激活的CD8αβ+T細胞上被瞬時(shí)誘導或在IEL上組成性地表達,其抑制作用既可以暫時(shí)性地降低功能親和力,減弱持續的免疫應答(圖1B),也可以永久性地增加有抗原刺激經(jīng)歷的T細胞重新激活所需的最小信號強度(圖1C)。盡管目前還不完全了解CD8αα是如何作為共抑制受體發(fā)揮作用的,但它與MHCI類(lèi)配體相互作用的能力以及它與TCR-CD3復合物的各種信號成分聯(lián)系的能力表明,CD8αα有可能在不同層次上干擾TCR介導的激活。
與CD8αβ連同TCR-CD3激活誘導的共內化不同,CD8αα的表達隨著(zhù)激活而增加,反映了CD8αα與TCR-CD3激活復合物之間的脫節關(guān)系,這與它被排除在脂質(zhì)筏之外的結果是一致的。脂質(zhì)筏外側CD8αα表達的增加與激活復合物功能親和力的降低相一致,暗示CD8αα被排除在脂質(zhì)筏之外可能是其抑制作用的關(guān)鍵。因此,CD8αα可能與非脂質(zhì)筏相關(guān)(non-lipid-raft-associated)的p56lck和LAT結合,從而將這些信號組分從CD8αβ和TCR-CD3激活復合物中分離出來(lái)(圖1)。盡管這是一個(gè)合理的假設,但CD8αβ輔助受體與p56lck和LAT有效結合能力的增強,表明還有其他機制與脂質(zhì)筏的排斥作用一起,共同有助于CD8αα的抑制作用。CD8αβ與TCR-CD3激活復合物的有效共定位取決于脂質(zhì)筏的完整性和更大的膜平臺組織。CD8αα的表達顯著(zhù)降低了CD8αβ和TCR-CD3的共定位和關(guān)聯(lián)性,這可能表明CD8αα主動(dòng)破壞了脂質(zhì)筏的完整性,從而破壞了CD8αβ與TCR的最佳關(guān)聯(lián)性。脂質(zhì)筏破壞作為一種干擾TCR-CD3激活的機制已經(jīng)被報道過(guò),在包括CTLA-4在內的其它TCR抑制因子。與CD8αα-CD8αβ輔助受體對類(lèi)似,CTLA-4與它的競爭者CD28一樣與B7配體相結合,但顯示出相反的功能:CD28作為T(mén)CR的共刺激受體,而CTLA-4介導對TCR的抑制活性。CTLA-4的誘導也與TCR信號強度有關(guān),綜合這些結果表明,脂質(zhì)筏破壞和對信號分子共定位的干擾可能是TCR輔助受體配對阻遏物的一個(gè)普遍特征。CD8αα的抑制活性并不限于CD8αβ,CD8αα介導的對雙陰性胸腺細胞的抑制作用,暗示其可以直接起到負調控TCR-CD3復合物的作用,而與傳統的TCR輔助受體無(wú)關(guān)(圖1C)。
CD8α的細胞質(zhì)尾部不包含任何典型的抑制性受體免疫受體酪氨酸抑制基序(ITIMs)。相反,CD8αα可以積極地與抑制性分子結合,就像所報道過(guò)的LAG-3那樣,LAG-3是一種與TCR輔助受體CD4有密切同源性的TCR激活誘導的抑制物。與CD8αα-CD8αβ受體對類(lèi)似,LAG-3與CD4共享MHC II類(lèi)配體,但LAG-3會(huì )通過(guò)積極招募細胞內抑制分子,部分負調控TCR-CD3的激活。雖然還沒(méi)有發(fā)現直接與CD8α胞質(zhì)尾部結合的特異性抑制分子,但我們很容易能夠推測,B細胞激活的連接物L(fēng)AT2,一種LAT的同源物,在幼稚T細胞中不表達,但在激活的T細胞上會(huì )被瞬時(shí)誘導表達且在CD8αα+IEL中大量表達,可能作為CD8αα的一種抑制因子。盡管LAT2在與包括FcεRIγ在內的含ITAM的激活受體結合時(shí)也會(huì )被磷酸化,但它可以主動(dòng)與LAT競爭,負向調節其在T細胞中的活性。值得注意的是,在激活的T細胞上瞬時(shí)誘導表達的CD8αα和LAT2以及它們在CD8αα+ IEL上組成性表達,也與瞬時(shí)或組成性表達的FcεRIγ相吻合(圖1C)。在T細胞中,FcεRIγ可以參與形成CD3復合物,可以取代CD3ζ二聚體或與CD3ζ形成異質(zhì)二聚體。每條CD3ζ鏈中存在的三個(gè)ITAM基序,而每個(gè)FcεRIγ單元只有一個(gè)ITAM,因此,CD3磷酸化位點(diǎn)的減少,加上LAT2對LAT的抵消作用和CD8αα的存在,這些抑制機制協(xié)同作用,提高了T細胞有效激活的閾值。膜結合的CD8αα負向調節輔助受體依賴(lài)性或輔助受體非依賴(lài)性TCR的激活,與其胞外結構域是否與抗原呈遞的MHC I類(lèi)分子有效結合無(wú)關(guān)。這與可溶性CD8αα(sCD8αα)分子或封阻MHC和CD8αβ輔助受體之間相互作用的CD8α衍生肽的作用形成鮮明的對比。sCD8αα的干擾導致CD3ζ的酪氨酸磷酸化受到抑制,并表明sCD8αα的阻斷針對的是激活的最早階段,這與p56lck激活的抑制相一致。sCD8αα的抑制作用不影響DN T細胞上表達的高親和力TCR,意味著(zhù)sCD8αα的抑制機制專(zhuān)門(mén)針對的是需要CD8αβ共受體功能才能激活的低親和力TCR。用阻斷CD8αβ與MHC配體相互作用的CD8抗體也觀(guān)察到類(lèi)似的抑制作用。sCD8αα的抑制作用是非常有效的,只有少數CD8αβ-MHC I類(lèi)的相互作用需要被sCD8αα阻斷,以防止T細胞的激活。sCD8αα產(chǎn)生于選擇性剪接的mRNA,其中編碼跨膜結構域的外顯子已被刪除。sCD8αα在人類(lèi)中自然發(fā)生,與多種疾病的晚期相關(guān),包括T細胞白血病、類(lèi)風(fēng)濕性關(guān)節炎、多發(fā)性硬化癥、系統性紅斑狼瘡和HIV感染。在小鼠中,一種可選擇性剪接形式的CD8α也同樣被報道,但與人類(lèi)的sCD8αα不同,小鼠CD8α mRNA保留了跨膜結構域,形成了一種尾部缺失的膜結合CD8α'分子。盡管所有的T細胞都轉錄和翻譯CD8α',但只有未成熟的胸腺細胞在其細胞表面表達CD8α'β異二聚體,而成熟的T細胞則在細胞內保留末端唾液酸化的含CD8α'的復合體。膜結合的CD8α'缺乏與p56lck結合的能力,因此,即使它有一個(gè)完整的細胞外結構域,可以與MHC I類(lèi)分子相互作用,但它不能作為T(mén)CR共受體亞單位發(fā)揮作用。值得注意的是,在低等脊椎動(dòng)物中表達的CD8α的祖先形式也保留了原型CD8 Ig樣外結構域以及鉸鏈和跨膜結構域,而它在細胞質(zhì)部分缺乏p56lck一致性結合基序,暗示這兩個(gè)物理上相連的功能單元可能是分別進(jìn)化而來(lái)的。
結論:由于與CD8αβ輔助受體在結構上有驚人的同源性,CD8αα的外表具有欺騙性,人們對它有很多假設。這種誤解不僅導致了對CD8αα功能的錯誤解釋?zhuān)疫€對表達CD8αα的T細胞的個(gè)體發(fā)生和功能產(chǎn)生了深刻的混淆和誤解。CD8αα的存在已被用作非常規IEL及其胸腺前身細胞,以及常規的CD8αβ+記憶前體細胞的標志。若假設該分子是多余的、無(wú)效的輔助受體時(shí),T細胞上CD8αα的表達僅僅是一個(gè)無(wú)關(guān)緊要的標記。當CD8αα被認為是在未成熟的胸腺細胞或初級效應細胞上誘導的強有力的共抑制受體時(shí),或在腸道中有抗原經(jīng)歷的成熟細胞上重新表達時(shí),CD8αα作為激活和分化的關(guān)鍵調控者的重要性就毋庸置疑了。總之,考慮CD8αα的抑制功能是非常重要的,包括對該分子今后的分析,對CD8αα在T細胞分化過(guò)程中作用的重新解釋?zhuān)约皩@表達CD8αα的T細胞的個(gè)體發(fā)育和功能爭議的重新解釋。
1. Cheroutre H, Lambolez F: Doubting the TCR coreceptor function of CD8alphaalpha. Immunity 2008, 28(2):149-159.